编者按:毒素-抗毒素系统(TA)与结核病耐药和持留菌的产生密切相关,但目前关于TA系统对结核分枝杆菌生理和致病机制的影响尚不完全清楚。本期“深三院结核之窗”栏目中,卢水华教授团队分享了一项发表于《自然-通讯》(Nat Commun)的研究,解释了MenT3/MenT4毒素蛋白对结核分枝杆菌毒力的影响和机制,并探究其作为结核病疫苗的可行性。
研究简介
结核分枝杆菌的基因组编码了大量的毒素-抗毒素系统。在本研究中,属于 TA 系统 MenAT 亚家族的 MenT3 和 MenT4 毒素已被功能表征。我们证明,这些毒素的异位表达抑制了细菌的生长,而这在其同源抗毒素的共同表达中得到了挽救。在这里,我们发现 menT3 和 menT4 的同时缺失导致豚鼠和小鼠在暴露于氧化应激时结核分枝杆菌的易感性增强和生长减弱。我们观察到,与亲本菌株相比,在 ΔmenT4ΔT3 的对数期培养物中,编码细胞内生长所必需或必需的蛋白质的转录本的表达降低。此外,相对于亲本品系感染的小鼠,ΔmenT4ΔT3感染小鼠的肺组织中参与有效细菌清除的蛋白质的转录水平增加。我们发现,用 ΔmenT4ΔT3 对小鼠和豚鼠进行免疫接种可显着预防结核分枝杆菌感染。值得注意的是,用 ΔmenT4ΔT3 免疫小鼠导致抗原特异性 TH1细胞反应增加,并激活了更多记忆T细胞反应。我们得出的结论是,MenT3 和 MenT4 对结核分枝杆菌的致病性很重要,menT3/menT4敲除菌株有可能作为候选疫苗进行进一步探索。
MenT3 和 MenT4 的异位表达导致生长抑制
本实验研究了MenT3和MenT4的过表达对大肠杆菌和结核分枝杆菌生长的影响。结果显示,MenT3和MenT4的过表达显著抑制大肠杆菌生长,而特定突变(如G62、K189、D208和G51、D71)则导致毒性丧失。抗毒素MenA3和MenA4的共表达能够恢复生长抑制,而非同源抗毒素无效。此外,MenT3和MenT4也抑制了结核分枝杆菌的生长,诱导后4天生长减少约5倍,7天后分别增加到28倍和15倍的生长缺陷。
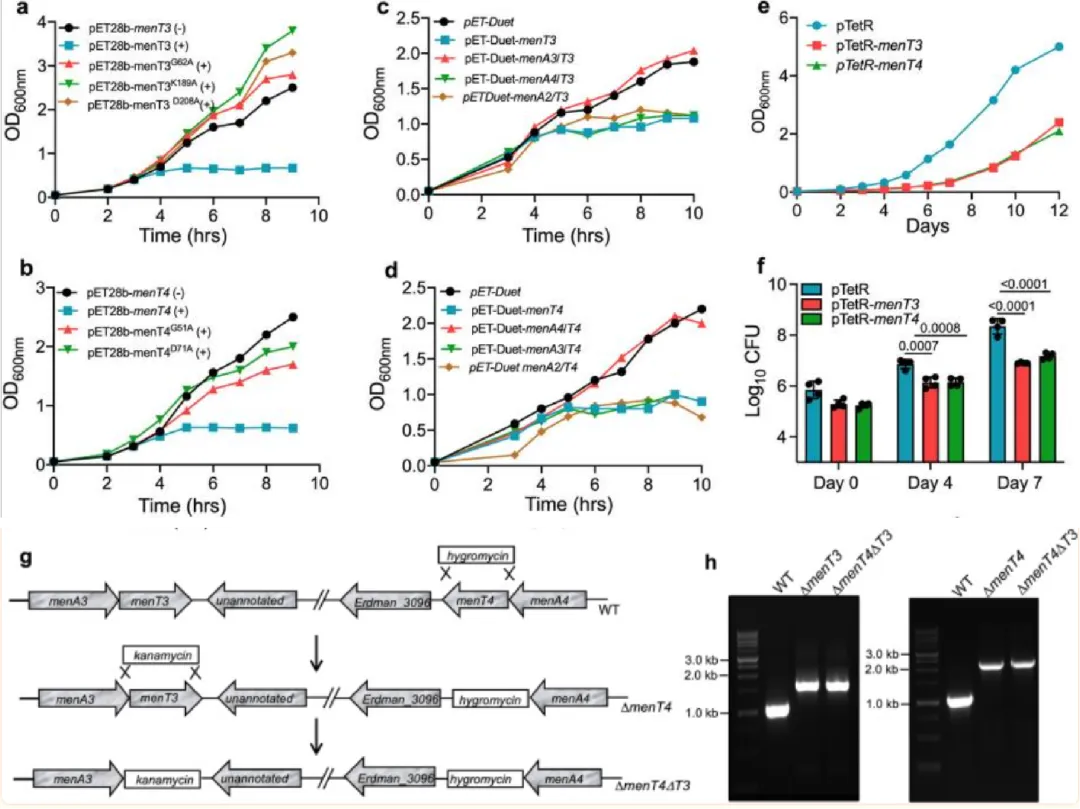
图1.属于结核分枝杆菌 TA 系统 MenAT 亚家族的 MenT3 和 MenT4 毒素的功能表征。a-f MenT3 和 MenT4 的过表达抑制大肠杆菌和结核分枝杆菌的生长。a, b 这些面板显示了大肠杆菌 Bl-21 (pLysS, λDE3) 菌株的生长模式,这些菌株携带表达野生型或突变型 MenT3 (a) 或野生型或突变型 MenT4 (b) 蛋白的 pET28b 衍生物,无论是否存在诱导剂。c, d 这些面板描绘了大肠杆菌 BL21 (pLysS, λDE3) 菌株的生长模式,这些菌株携带过表达 MenT3 (c) 或 MenT4 (d) 的 pET-Duet 结构,可以单独使用或与其同源或非同源抗毒素一起表达。通过测量OD来确定各种菌株的生长情况600纳米.这些面板中显示的数据代表了两个独立的实验。e, f 结核分枝杆菌H的生长模式37这些面板显示了含有表达 MenT3 或 MenT4 的 pTetR 衍生物的 Rv。重组菌株的生长是通过测量 OD 来确定的600纳米 (e) 或细菌计数 (f)。(e)中显示的数据代表了两个独立的实验。(f)中显示的数据是对数的平均值±标准差10CFU来自两个独立实验,每个实验都使用重复培养物进行。图表上描绘的 p 值使用单因素方差分析进行评估。g, h结核分枝杆菌ΔmenT4ΔT3菌株的构建。g 显示了结核分枝杆菌 Erdman 的亲本、ΔmenT4 和 ΔmenT4ΔT3 菌株中 menT3 和 menT4 位点的示意图。将结核分枝杆菌ΔmenT4菌株的潮霉素抗性基因替换了menT4的开放阅读框。在双突变株ΔmenT4ΔT3中,menT3和menT4的开放阅读框分别被卡那霉素和潮霉素抗性基因取代。h 使用基因特异性引物的PCR证实了menT3和menT4在各自的单突变株和双突变株中分别用卡那霉素和潮霉素抗性基因替代。源数据以源数据文件的形式提供。
menT3 和 menT4 的缺失增加了结核分枝杆菌对氧化应激的易感性,并损害豚鼠中结核分枝杆菌的毒力
在液体培养中,野生型与ΔmenT4ΔT3菌株的细菌计数相似,但在氧化应激下,双突变株显示出显著的敏感性,24小时后敏感性高出约8.5倍,72小时后高出约225倍。与menT3互补后,双突变株的生长缺陷得以恢复,但与menT4互补则无效。qPCR结果表明,单互补菌株中menT3和menT4的转录水平恢复。此外,ΔmenT4ΔT3菌株在氧化应激、亚硝化、营养和酸性胁迫下的存活率与亲本菌株相当。对THP-1巨噬细胞的感染实验显示,所有菌株在巨噬细胞内的生长模式相似。综上所述,MenT3和MenT4在体外生长中是互补的,缺失这两个基因会增加结核分枝杆菌对氧化应激的敏感性。
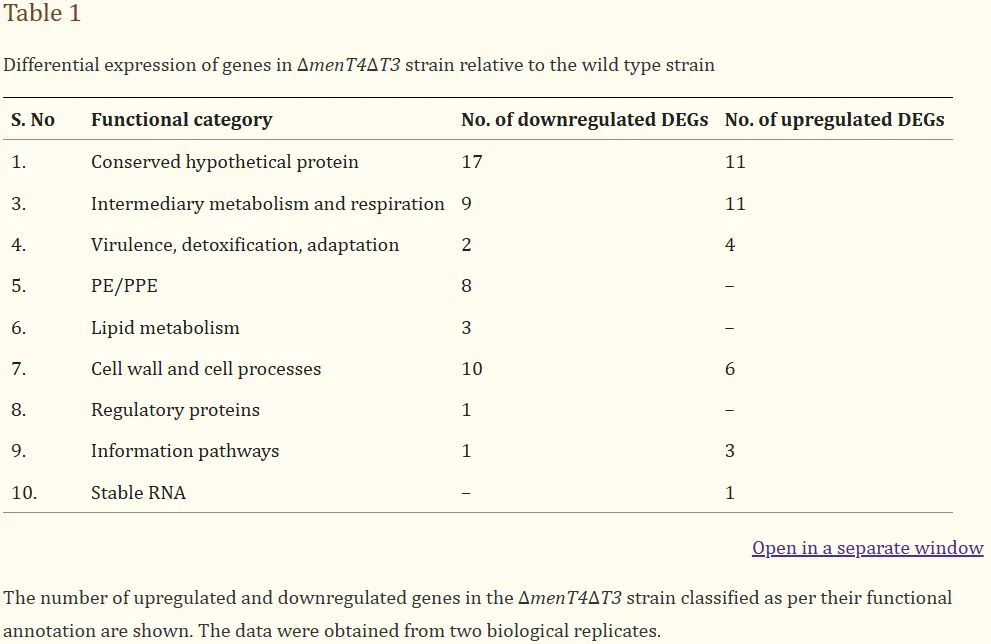
通过比较气溶胶感染的野生型与ΔmenT4ΔT3双突变株的生长,发现感染ΔmenT4ΔT3的豚鼠在感染后4周和8周的肺杆菌负荷分别降低了约6.6倍和950倍,脾脏细菌负荷则减少了约117倍和3750倍。此外,H&E染色结果显示,感染野生型菌株的豚鼠肺部组织存在明显的细胞浸润和炎症,而ΔmenT4ΔT3感染的豚鼠则保持了完整的肺结构和较大的肺泡空间。总肉芽肿评分在ΔmenT4ΔT3感染组显著降低了约10倍。综上所述,MenT3和MenT4的同时缺失显著减弱了豚鼠结核分枝杆菌的生长和致病性。
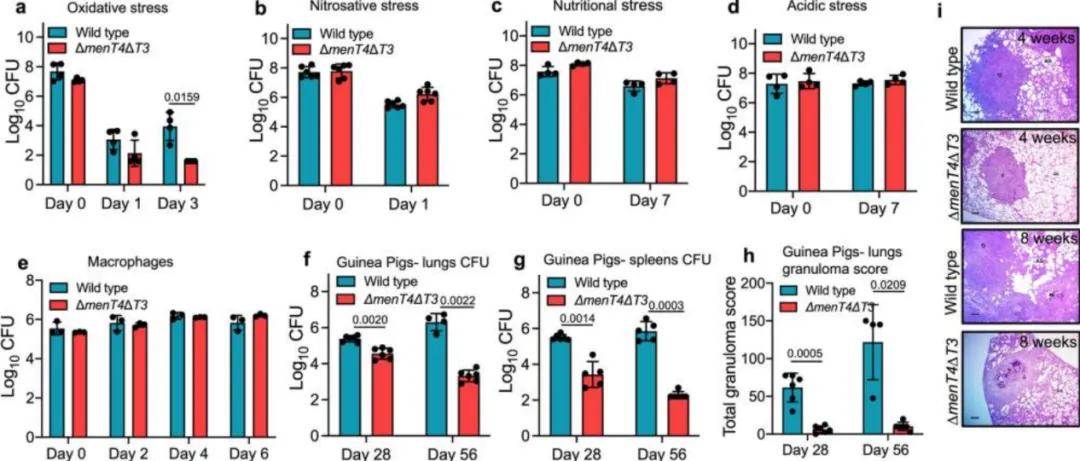
图2. MenT3 和 MenT4 对于结核分枝杆菌在氧化应激中存活并在豚鼠中建立感染是累积必需的。比较了野生型和 ΔmenT4ΔT3 暴露于氧化 (a) 或亚硝化 (b) 或营养 (c) 或酸性 (d) 胁迫后的生长情况。这些面板中显示的数据是对数的平均值± SD10CFU来自两个(a、c、d)或三个(b)独立实验,每个实验都用重复的培养物进行。e THP-1巨噬细胞感染多种菌株,测定不同时间点的细胞内细菌数量。此面板中显示的数据是对数的平均值± SD10从一式三份的孔中获得的CFU,代表了以一式两份或一式三份进行的两次独立实验。这些面板中显示的数据以对数的平均值± SD 表示10感染后 4 周和 8 周感染野生型或 ΔmenT4ΔT3 菌株的豚鼠(Duncan Hartley 品系)的肺 (f) 和脾脏 (g) 中的 CFU。这些面板中显示的数据以对数的平均值± SD 表示10从 6 只动物获得的 CFU(除了 (f) 第 8 周,野生型 n = 4 和 (g) 第 4 周,ΔmenT4ΔT3 n = 5 和第 8 周,野生型 n = 5)。显示的 4 周时间点的数据代表了两个独立的实验。显示的 8 周时间点的数据是从单个实验中获得的。h 该面板中显示的数据是感染野生型或 ΔmenT4 Δ T3菌株的结核分枝杆菌野生型或 ΔmenT4 Δ T3 菌株的 H&E 染色切片中总肉芽肿评分的平均值± SD。显示的数据是从单个实验的 6 只动物中获得的(第 8 周除外,野生型 n = 4)。i 该面板显示了感染后 4 周或 8 周感染野生型或 ΔmenT4ΔT3 菌株的豚鼠肺组织 H&E 染色切片的代表性图像。比例尺,100μm。图表上描绘的 P 值是使用双尾配对 t 检验评估的。源数据以源数据文件的形式提供。
menT3 和 menT4 的缺失改变了结核分枝杆菌的转录谱
本实验比较了野生型与ΔmenT4ΔT3双突变株在对数期培养物中的转录谱,以探讨MenT3和MenT4缺失对结核分枝杆菌的影响。结果显示,相对于亲本菌株,ΔmenT4ΔT3菌株中编码36种和51种蛋白质的转录本分别增加和减少33%和23%的差异表达基因(DEGs)涉及保守假设蛋白质或代谢与呼吸相关的蛋白质。特别地,II型TA系统的毒素(如vapC24、vapC27、vapC36和vapC38)在ΔmenT4ΔT3中表达上调。此外,参与结核分枝杆菌适应宿主的蛋白质转录本(如Erdman_0097和Erdman_0100)在ΔmenT4ΔT3中显著降低,提示其在低氧条件下的调节功能受损。也观察到与宿主干扰素-γ相互作用的蛋白质的转录水平下降。尽管抗生素标记可能影响邻近基因表达,研究显示MenT3和MenT4相邻基因的转录水平在野生型和突变株中相似。此外,非极性和极性脂质的相对水平在不同菌株中无显著差异,排除了脂质损失的可能性。综上所述,ΔmenT4ΔT3菌株的减毒表型主要与结核分枝杆菌应激适应或毒力相关蛋白质的转录本水平降低有关。
menT3 和 menT4 的缺失导致小鼠结核分枝杆菌生长减弱
本实验比较了感染后4周和8周气溶胶感染的Balb/c小鼠中野生型与ΔmenT4ΔT3双突变株的生长情况。结果显示,ΔmenT4ΔT3感染的小鼠肺和脾脏杆菌负荷在感染后4周分别降低了10倍和14倍,8周后分别降低7倍和13倍,表明MenT3和MenT4增强了结核分枝杆菌在宿主中的感染能力。进一步的RNA-seq分析揭示,感染ΔmenT4ΔT3菌株的小鼠中,相比未感染小鼠,312个和119个基因的转录水平分别增加或减少。与亲本菌株感染的小鼠相比,ΔmenT4ΔT3感染小鼠的肺组织中编码参与钙信号传导的蛋白质的基因转录水平增加。此外,感染突变株的小鼠中,涉及自噬和凋亡的相关基因(如Bub1、Phf23和Atg16l1)的转录本表达差异明显,提示自噬机制可能受到影响。免疫反应相关的转录本(如IL-21、CCL1、CXCL5等)在ΔmenT4ΔT3感染小鼠中显著降低,表明其炎症反应减弱。与此同时,编码参与巨噬细胞、中性粒细胞或T细胞活化的基因转录水平有所增加,提示可能增强了对细菌的免疫反应。综上所述,ΔmenT4ΔT3感染小鼠的减毒表型可能与钙稳态、凋亡、自噬相关蛋白表达的增加,以及与炎症反应相关转录本表达的减少有关。
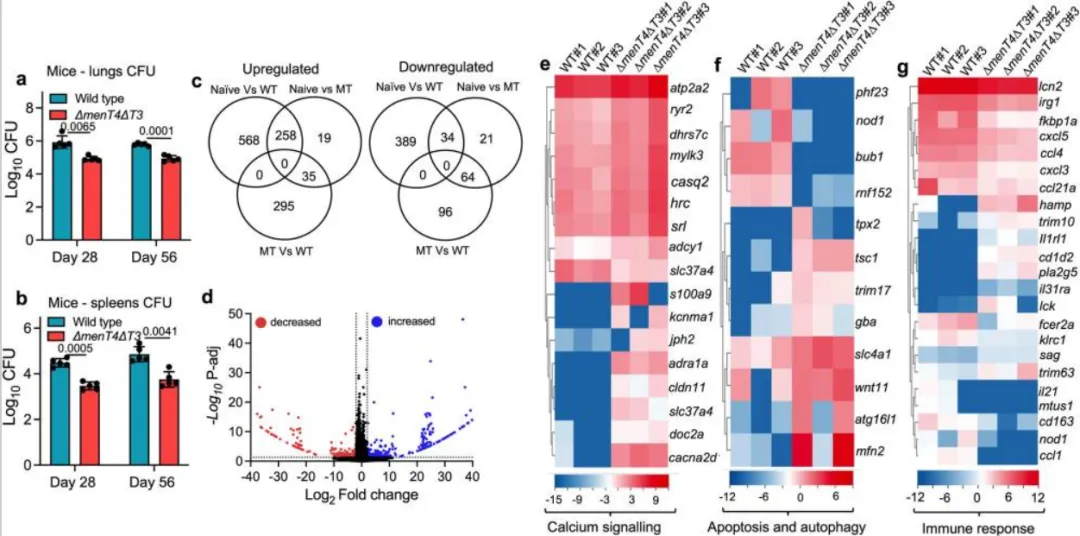
图3. 感染后 4 周感染野生型或 ΔmenT4ΔT3 菌株的小鼠肺组织的全局转录变化。a,b menT3 和 menT4 的缺失可减弱小鼠肺和脾脏中结核分枝杆菌的生长。 这些面板中显示的数据是对数的平均值± SD10感染后 4 周和 8 周从 5 只动物获得的感染 Balb/c 小鼠的肺 (a) 或脾脏 (b) 中的 CFU。这些面板中显示的 4 周时间点的数据代表了两个独立的实验。显示的 8 周时间点的数据是从单个实验中获得的。图表上描绘的 P 值是使用双尾配对 t 检验评估的。来自未感染或感染野生型或 ΔmenT4ΔT3 菌株的小鼠肺组织的 c-g 宿主转录谱。c 维恩图描绘了感染后 4 周从未感染或感染亲本或 ΔmenT4ΔT3 菌株的小鼠肺组织获得的表达谱的相关性。d 火山图比较了感染后 4 周从感染野生型或 ΔmenT4ΔT3 菌株的小鼠肺组织获得的转录谱。表达增加或减少的转录本分别显示为蓝点或红点。黑点表示保持不变的转录本,并且在这两组之间没有统计学上的差异。例如-g:热图显示了感染后 4 周感染野生型或 ΔmenT4ΔT3 菌株的小鼠中具有不同表达的转录本。在感染这些菌株的小鼠中具有差异表达的转录本参与钙信号传导 (e) 或凋亡/自噬 (f) 或免疫反应 (g)。热图中的颜色强度表示对数2归一化表达式计数的值。(c-g)中显示的数据是从三个独立的生物学重复中获得的。源数据以源数据文件的形式提供。
用 ∆menT4∆T3 菌株免疫可保护小鼠和豚鼠免受结核分枝杆菌攻击
本实验评估了ΔmenT4ΔT3菌株在C57BL/6小鼠和Duncan Hartley豚鼠中对结核分枝杆菌感染的保护作用。结果显示,免疫接种ΔmenT4ΔT3的小鼠在感染后6周,肺和脾脏中的免疫杆菌数量低于检测限。感染后4周,ΔmenT4ΔT3免疫小鼠的肺和脾脏杆菌负荷分别减少了约7.5倍和5.0倍,且在感染后10周,仍保持显著的降低(肺和脾脏负荷分别减少4.5倍和18.5倍)。与幼稚小鼠相比,BCG免疫在肺和脾脏中的细菌计数也显著减少。
在豚鼠中,BCG和ΔmenT4ΔT3免疫后,肺和脾脏的细菌载量在免疫6周后同样低于检测限。接种ΔmenT4ΔT3的豚鼠在感染后4周的肺部细菌数量减少约70倍,而BCG疫苗的豚鼠仅减少6倍。感染后8周,ΔmenT4ΔT3免疫的豚鼠肺杆菌负荷降低约27倍,保护效果较BCG提高了约2.5倍,尽管统计学意义不显著。
组织病理学分析显示,未接种疫苗的豚鼠在感染后8周的总肉芽肿评分为约50,而接种BCG的动物降低至29,接种ΔmenT4ΔT3的豚鼠则显著降低至8.5和0.4,表明其在减少病理损伤方面的有效性。整体而言,ΔmenT4ΔT3的免疫接种在小鼠和豚鼠中均显示出显著的保护作用,减少了细菌负荷和组织损伤。
图4用 ΔmenT4ΔT3 对小鼠和豚鼠进行免疫可预防结核分枝杆菌攻击。6-8 周龄雌性 C57BL/6 小鼠皮下注射 5 × 105牛支原体 BCG 或 ΔmenT4ΔT3 的 CFU。在免疫后 10 周,用结核分枝杆菌攻击动物,并在感染后 4 周和 10 周进行细菌计数。b–e 这些面板中显示的数据是对数的平均值± SD10幼稚的肺(b,d)和脾脏(c,e)中的CFU或BCG或ΔmenT4ΔT3免疫小鼠在感染后4周(b,c)和10周(d,e)感染后使用结核分枝杆菌攻击后。 这些面板中显示的数据是对数的平均值± SD10从单一实验中获得的 5 只动物的 CFU(除了在 (e) 第 10 周,ΔmenT4ΔT3 n = 4)。f 6-8周龄的雌性豚鼠(Duncan Hartley品系)皮内免疫1×105牛支原体 BCG 或 ΔmenT4ΔT3 的 CFU。免疫后 10 周,对动物进行结核分枝杆菌攻击,并在感染后 4 周和 8 周进行细菌计数和组织病理学分析。幼稚或 BCG 或 ΔmenT4ΔT3 免疫豚鼠肺部的细菌负荷在攻击后 4 (g) 和 8 周 (h) 用结核分枝杆菌攻击后测定。显示的数据是对数的平均值± SD10从单一实验中获得的 6 只动物的 CFU(除了 (g) 第 4 周,幼稚和 BCG n = 7,以及 (h) 第 8 周,BCG n = 8)。该面板中显示的数据是在攻击后 4 周 (i) 和 8 周 (j) 从幼稚或 BCG 或 ΔmenT4ΔT3 免疫豚鼠的 H&E 染色肺切片获得的总肉芽肿评分的平均值± SD。显示的数据是从单个实验的6只动物中获得的(除了(i)第4周,幼稚和BCG n = 7和(j)第8周,BCG n = 8)。k 该面板显示了初治或免疫豚鼠感染结核分枝杆菌 4 周或 8 周后肺组织的 H&E 染色切片的代表性图像。比例尺,100μm。图表上描绘的 p 值使用单因素方差分析进行评估。源数据以源数据文件的形式提供。
用 ∆menT4∆T3 免疫小鼠诱导 TH1细胞反应和激活记忆 T 细胞反应
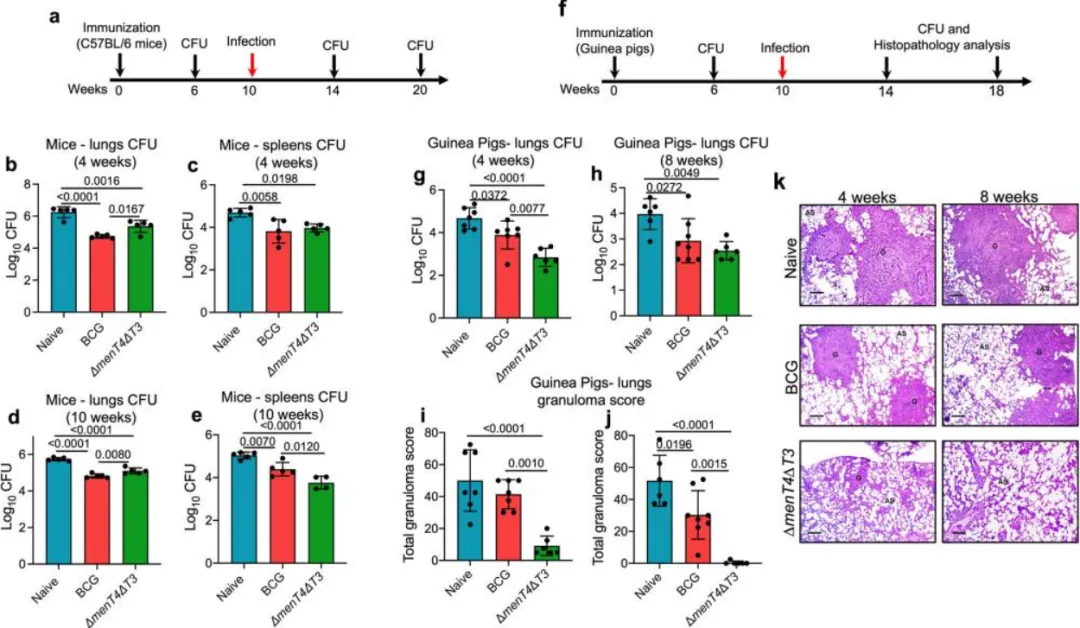
本研究评估了ΔmenT4ΔT3免疫对C57BL/6小鼠的保护作用及其免疫相关性。结果显示,与幼稚小鼠相比,ΔmenT4ΔT3免疫的小鼠中抗原特异性TH1细胞反应显著增强,IFN-γ阳性CD4 T细胞的频率增加约270%和190%,而IFN-γ阳性CD8 T细胞的频率增加300%-450%。此外,T-bet的表达水平在ΔmenT4ΔT3免疫小鼠中也显著提高,表明其对TH1反应的影响。
在细胞因子水平上,PPD刺激后,ΔmenT4ΔT3免疫小鼠的IFN-γ和TNF-α水平显著高于幼稚小鼠,IL-2水平亦有所增加,而IL-17A水平变化不显著。此外,免疫并未导致FoxP3阳性调节性T细胞的显著变化,提示ΔmenT4ΔT3免疫的保护作用可能独立于Treg细胞。
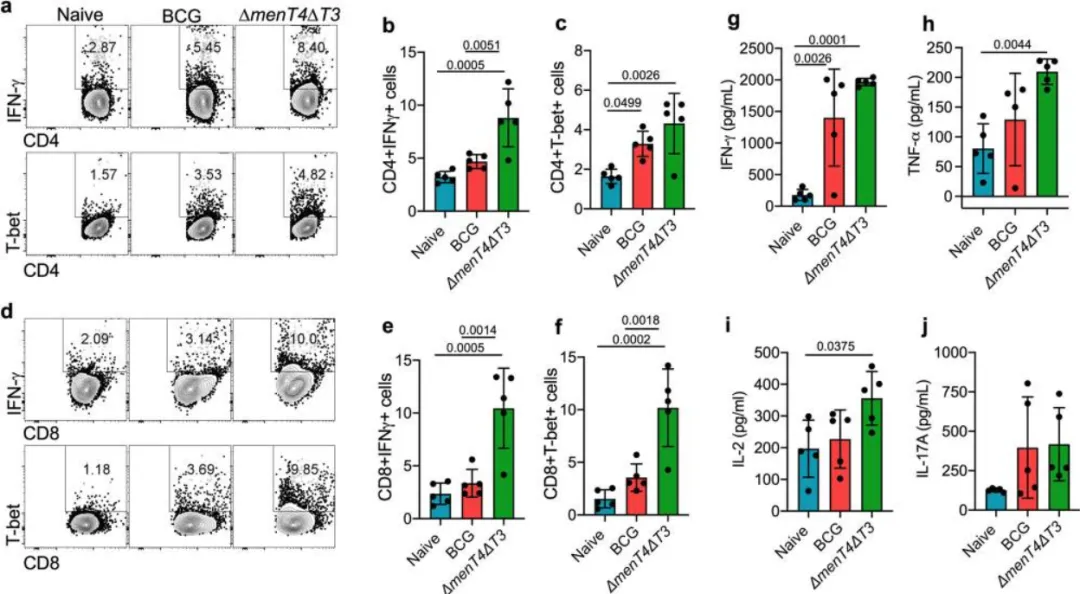
进一步分析显示,ΔmenT4ΔT3免疫小鼠的记忆T细胞反应增强,激活的记忆CD4和CD8 T细胞频率分别增加2.70倍和3.80倍,相比BCG免疫小鼠的提升也显著。虽然效应记忆CD4 T细胞的比例在ΔmenT4ΔT3免疫小鼠中有所下降,但总体效应和记忆CD8 T细胞的频率相当。
综上所述,ΔmenT4ΔT3免疫能有效增强抗原特异性TH1细胞反应和记忆T细胞的激活,可能为对结核分枝杆菌感染提供更强的保护。
图5.用 ΔmenT4ΔT3 免疫小鼠导致抗原特异性 T 增加H1-回应。在免疫后 6 周处死用生理盐水或 BCG 或 ΔmenT4ΔT3 免疫的 a-f C57BL/6 小鼠,并在 PPD 刺激的脾细胞中测量细胞内细胞因子。a 代表性FACS图,描绘了初治或BCG或ΔmenT4 Δ T3免疫小鼠脾脏中CD4 IFN-γ和CD4 T-bet T细胞的百分比频率。这些面板显示了初治或 BCG 或 ΔmenT4ΔT3 免疫小鼠脾脏中百分比频率 CD4 IFN-γ (b) 和 CD4 T-bet (c) 的平均 ± SD。d 代表性FACS图,描绘了幼稚或BCG或ΔmenT4 Δ T3免疫小鼠脾脏中CD8 IFN-γ和CD8 T-bet T细胞的百分比频率。这些面板显示了幼稚或 BCG 或 ΔmenT4ΔT3 免疫小鼠脾脏中 CD8 IFN-γ (e) 和 CD8 T-bet (f) T 细胞的百分比频率。(b、c、e、f)中显示的数据是从单一实验中获得的五只动物的T细胞比例的平均值±标准差。采用ELISA法测定PPD刺激的脾细胞培养上清液中分泌的IFN-γ(g)、TNF-α(h)、IL-2(i)、IL-17A(j)水平。显示的数据是从单一实验中获得的五只动物的PPD刺激的脾细胞中细胞因子水平的平均值±SD((h)除外,BCG n = 4)。图表上描绘的 p 值使用单因素方差分析进行评估。源数据以源数据文件的形式提供。
图6.用ΔmenT4ΔT3 免疫小鼠增加了激活记忆 T 细胞反应的扩展。代表性 FACS 图显示 CD4 激活记忆 T 的百分比频率+H细胞(CD4、CD44、CD69、a)和 CD8 激活记忆 T++++C幼稚或BCG或ΔmenT4ΔT3免疫C57BL / 6小鼠脾脏中的细胞(CD8,CD44,CD69,b)。 这些面板显示了激活内存 CD4 T 的百分比频率++++H细胞 (c) 和 CD8 T+C幼稚或BCG或ΔmenT4ΔT3免疫小鼠的脾脏中的细胞(d)。这些面板中显示的数据是从单次实验中获得的五只动物的细胞百分比频率的平均值±标准差。图表上描绘的 p 值使用单因素方差分析进行评估。源数据以源数据文件的形式提供。
研究点评
2022年,全球报告的新诊断结核病病例数达到750万。这是自1995年世界卫生组织开始全球结核病监测以来报告的最高数字,超过了2019年新冠肺炎大流行前的基线和710万例的历史峰值。尽管开发了许多有效的抗结核药物,但结核病仍然是一个主要的全球健康问题。一方面,结核病的常规抗生素治疗过程漫长,并伴有肝毒性和其他毒副作用,导致患者依从性差和潜在的治疗失败。另一方面,结核分枝杆菌对新旧抗结核药物都产生了耐药性,对耐药患者的治疗可能持续20多个月,成本很高。虽然卡介苗能够给新生儿提供一定的针对结核分枝杆菌感染的保护力,但保护力随着年龄增加而减弱,特别是目前没有一款疫苗能够有效地保护中老年人。此外,HIV-1和结核分枝杆菌的共同感染显著增加了活动性结核病的风险,使结核病的治疗更加复杂。这些挑战大大增加了结核病治疗的成本和负担,推动了结核病的全球传播。因此,仍然需要不断开发有效的抗结核药物,并开发新的结核病疫苗以保护所有易感人群。
毒素-抗毒素系统(TA系统)是原核生物普遍存在的小遗传元件,由两个基因组成,分别编码稳定表达的毒素蛋白和不稳定表达的抗毒素蛋白。作者之前的研究发现,MenT3 和 MenT4 与 IV 型 TA 系统的毒素具有序列同源性,属于核苷酸转移酶DUF1814家族。MenT毒素的过表达通过阻止氨酰化和tRNA充电来抑制细菌生长。在此研究基础上,作者进一步探索了MenT3和MenT4在结核分枝杆菌生理学和发病机制中的作用。研究结果表明,尽管 MenT3 和 MenT4 对于体外生长是可有可无的,但这些毒素以累积方式发挥作用,对于结核分枝杆菌在小鼠和豚鼠中建立疾病至关重要。MenT3 和 MenT4 的缺失会导致结核分枝杆菌毒性减弱,与应激和毒力蛋白相关基因的下调有关。将∆menT4∆T3缺失株感染小鼠后,小鼠肺部自噬和凋亡水平增加,炎症损伤相关基因显著下调,表明∆menT4∆T3缺失株能够诱导小鼠的抗感染免疫并减少肺部炎症损伤。研究还表明,用 ΔmenT4ΔT3 免疫能够在小鼠和豚鼠中提供对结核分枝杆菌的保护。在豚鼠模型中,ΔmenT4ΔT3的保护力要优于传统BCG疫苗。作者发现在 ∆menT4∆T3 免疫动物中观察到的保护作用最有可能与抗原特异性 TH1 细胞的增加以及激活的记忆免疫反应有关.本研究不仅有助于我们理解结核分枝杆菌的致病机制,还为基于 MenT3 和 MenT4 调节的毒力相关通路的灭活结核病活疫苗的开发提供了理论依据。
本研究依然存在一定局限性,即新疫苗对小鼠的保护力不足,需要采用其它动物模型进一步的验证。此外,疫苗的安全性评价和临床试验也有待进行。未来的研究包括(1)基于∆menT4∆T3的多等位基因突变菌株(如panCD、leuD、metX等)的去标记和构建,这些菌株将按照日内瓦共识进行安全性和有效性研究的评估(2)鉴定针对MenT3和MenT4蛋白的小分子抑制剂,以及验证(3)从感染肺组织获得的RNA序列数据的实验。
更多精彩内容,请扫码订阅“深三院结核之窗”专栏
卢水华 教授
教授,主任医师,二级教授,博士生导师
国家感染性疾病临床医学研究中心副主任,深圳市第三人民医院肺病医学部主任
担任:中华医学会结核病分会候任主委,世界卫生组织全球儿童和青少年结核病工作组成员,中国防痨协会学校与儿童结核病分会主委,上海市医学会结核病学分会荣誉主委,广东省医学会结核病学分会主委,第二届国家名医获得者,上海市十佳医师,国家十三五传染病重大专项负责人,国家自然基金重大课题负责人,国家卫健委流感医疗救治专家组成员,国家自然科学基金评审专家,国家药监局新药评审专家,国家药监局医疗器械评审专家,中华医学会医疗鉴定专家,多杂志副主编、编委及审稿专家
长期致力于TB的发病机制、疫苗与诊断技术开发、流行病学及新药临床试验等领域的研究:深入开展大规模人群队列研究,明确我国大学生人群Mtb感染率和BCG保护效率;构建和组织实施“结核感染免疫诊断分层解决方案”,牵头完成我国40年来首个结核I类新药(EC)的临床试验及上市;积极探索TB防控“关口前移”新策略,建立和推广结核潜伏感染早期筛查和精准干预体系;针对当前TB防治瓶颈问题,提出的“一体化综合防控策略”为“End TB”贡献了中国原创理论和技术产品体系。主持包括国家“十三五”传染病重大专项、重大新药创制项目、国家自然基金重大课题在内的国家、省部和市级科研项目11项,合计研究经费超7260万元。在NEJM、Lancet、PNAS等顶级期刊发表学术论文73篇,组织撰写指南与专家共识3篇,主编专著1部。相关成果获“中国防痨协会科学技术奖”一等奖和“上海医学科技奖”三等奖,多次被WHO指南引用
陈展鹏
医学硕士,实验员
国家感染性疾病临床医学研究中心,药效学评价平台实验员;汕头大学基础免疫学硕士,主要研究方向:抗结核药物及其作用机制,结核分枝杆菌感染与脂质代谢等;参与多项省级和国家自然基金项目
来源:《感染医线》
声 明
凡署名原创的文章版权属《感染医线》所有,欢迎分享、转载(开白可后台留言)。本文仅供医疗卫生专业人士了解最新医药资讯参考使用,不代表本平台观点。该等信息不能以任何方式取代专业的医疗指导,也不应被视为诊疗建议,如果该信息被用于资讯以外的目的,本站及作者不承担相关责任。